|
|
423 M-AM
Learning modulates indicators of relational complexity in the occipitoparietal lobe
|
Steven Phillips, Kazuhisa Niki
National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba, Japan
|
Relational
complexity theory explains cognitive difficulty in terms of the
relationships between task items (Halford, Wilson, and Phillips, 1998),
and a number of studies have reported increased prefrontal and parietal
activity with relational complexity (Christoff, et al., 1998; Kroger et
al, 2002; Phillips and Niki, 2003). Learning and expertise also
modulate the effect of complexity inherent in task design. For example,
complexity may be reduced by learning to recode two items as a single
item chunk. Alternatively, complexity may be increased by employing a
ternary relation between three items where a binary relation would
suffice. In previous work (Phillips and Niki, 2003), we identified a
bilateral region in the occipitoparietal lobe sensitive to changes in
relational complexity. Here, we report an extension to this work where
we investigated the effect of learning on sensitivity to differences in
relational complexity.
Eight Japanese subjects were scanned before, during and after a 2-week
training period during which they learned to recognized shape pairs.
For each scanning trial, subjects determined whether a probe pair
appeared in a previous three-pair study list. Lists were either AB CD
EF (low relational complexity), or AB AD CB (high relational
complexity) of novel or learned planar shapes. Study pairs were
presented serially for 2170 ms separated by a 2000 ms blank screen
(encoding phase). Following the encoding phase, there was an 8000 ms
blank screen delay period (retention phase) after which the probe pair
was presented for 3000 ms, followed by a “+” symbol (5000 ms) to
indicate the end of the trial (Figure 1). Scanning was performed on a
3.0-T MRI Scanner (GE 3T Signa) with EPI capability. 18 axial slices
(5.3 mm thick, interleaved) were set to cover the entire brain. A T2*
weighted gradient echo EPI was employed. The imaging parameters were
TR=2 sec, TE=30 ms, FA=70°, FOV=20x20 (64x64 mesh). Using SPM99, data
were preprocessed and fitted by a general linear model, where 11 (i.e.,
2 [low, high complexity] x 2 [novel, learned shapes] x 2 [encoding,
retention phase] + 3 [target, distractor, error trials]) event
conditions were defined (Phillips and Niki, in press, for details).
 Figure 1. Example shapes and trial sequence.
Figure 1. Example shapes and trial sequence.
Regions of interest (ROI) analysis, based on ROIs from our previous
study, revealed a significant interaction between complexity and
learning (P<.009) for a bilateral region with centers at Brodmann
co-ordinates [-22 -70 37] and [22 -78 41], which border on the
occipitoparietal sulcus (see Figure 2 for whole-brain contrast).
Further analysis suggested that subjects treated low complexity lists
as high complexity lists before training, possibly due to difficulty
maintaining distinct memory traces for novel shapes. These results
suggest that the occipitoparietal region provides a window into
effective relational complexity. That is, complexity as imposed on
cognitive brain processes, not as prespecified by task design.
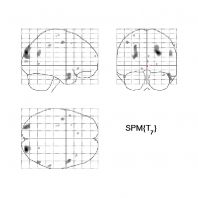 Figure 2. Complexity by learning contrast (p<.01, uncorrected).
Figure 2. Complexity by learning contrast (p<.01, uncorrected).
References
Christoff, et al. (1998). NeuroImage, 14(5), 1136-1149.
Kroger, et al. (2002). Cerebral Cortex, 12, 477-485.
Halford, Wilson, and Phillips (1998). Behavioral and Brain Sciences, 21(6), 803-831.
Phillips and Niki (2003). NeuroImage, 20(2), 1226-1235.
Phillips and Niki (in press). NeuroImage.
|
|

